
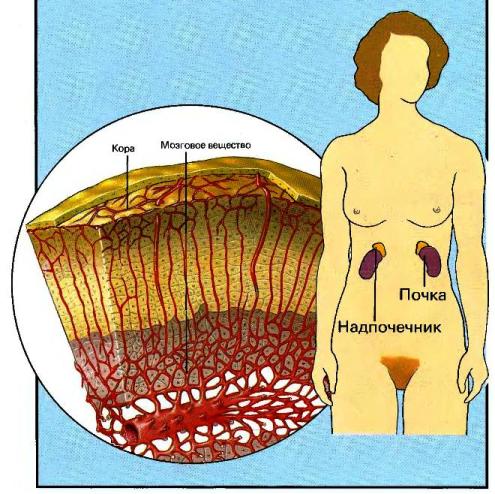
Строение надпочечников и синтез гормонов
Надпочечники состоят из двух видов эндокринной ткани — мозгового и коркового вещества. Хромаффинные клетки мозгового вещества имеют нейроэктодермальное происхождение, тогда как клетки коры — мезодермальное. Мезодермальные клетки участвуют также в развитии половых желез. Надпочечники и половые железы обладают рядом общих ферментов, необходимых для синтеза стероидов, врожденные дефекты стероидогенеза в одной ткани могут присутствовать и в другой.
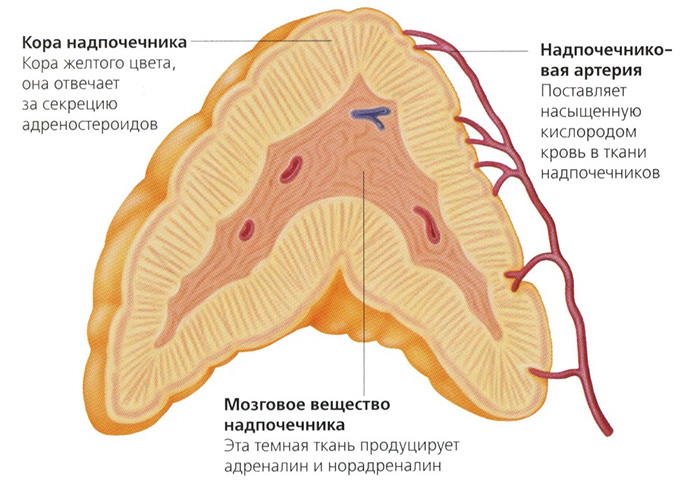
В коре надпочечников различают три зоны:
-
клубочковую (наружную, расположенную сразу под капсулой);
-
пучковую (промежуточную);
-
сетчатую (внутреннюю, окружающую мозговое вещество).
Самая крупная — пучковая зона, она занимает примерно 3/4 всего объема коркового вещества. На долю клубочковой зоны приходится около 15 % этого объема, а на долю сетчатой — примерно 10%. Для клеток клубочковой зоны характерны небольшие размеры, меньшее соотношение между объемами цитоплазмы и ядер, среднее количество липидных включений и меньшее число ядер с плотным хроматином, чем в клетках двух других зон. Клетки пучковой зоны крупные, и цитоплазма занимает в них большее пространство, чем ядро. Обилие липидных включений придает цитоплазме пенистый вид с большим числом вакуолей. Эти клетки располагаются в форме радиальных тяжей. Неравномерно расположенные клетки сетчатой зоны образуют между собой анастомозы. Соотношение цитоплазмы и ядра в них среднее, а количество липидных включений в плотной цитоплазме относительно мало.
В клубочковой зоне синтезируется альдостерон (наиболее активный природный минералокортикоид), в пучковой зоне — кортизол (наиболее активный природный глюкокортикоид), а в пучковой и сетчатой — надпочечниковые андрогены.
Мозговое вещество надпочечников состоит в основном из нейроэндокринных (хромаффинных) и глиальных (поддерживающих) клеток. В нем присутствует также некоторое количество соединительной ткани и сосудистых клеток. Нейроэндокринные клетки имеют полиэдрическую форму и содержат обильную цитоплазму с мелкими бледно окрашивающимися ядрами. При электронной микроскопии в их цитоплазме обнаруживается множество секреторных гранул, в которых содержатся катехоламины. В глиальных клетках меньше цитоплазмы, а ядра более базофильны.
Закладка надпочечников у плода различима уже на 3-4-й неделе внутриутробного периода. Она располагается сразу над развивающейся первичной почкой. На 5-6-й неделе клетки полового гребня превращаются в стероидогенные клетки половых желез и коры надпочечников. Первые мигрируют в каудальном направлении, вторые — в забрюшинное пространство. На 6-8-й неделе надпочечники быстро увеличиваются. Клетки внутренней коры дифференцируются, образуя зародышевую зону, а наружный субкапсулярный ободок остается в виде дефинитивной зоны. В это время в закладку коры надпочечников проникают элементы симпатической нервной системы, которые дифференцируются в хромаффинные клетки, синтезирующие и запасающие катехоламины.
Позднее в этих клетках появляется катехол-О-метилтрансфераза — фермент, превращающий норадреналин в адреналин. К концу 8-й недели внутриутробной жизни, окруженные капсулой надпочечники, вступают в контакт с верхними полюсами почек. В течение 9-12-й недели клетки зародышевой зоны активно вырабатывают стероиды. У 2-месячного плода надпочечники крупнее почек, но с 4-го месяца почки начинают быстро расти, становясь к концу 6-го месяца в 2 раза больше надпочечников. У доношенных новорожденных надпочечники по размеру в 3 раза меньше почек, общая масса обеих желез составляет 7-9 г. На долю внутренней зародышевой коры при рождении приходится примерно 80 % массы железы, а на долю наружной («истинной») коры — 20%. В первые дни постнатальной жизни зародышевая кора начинает сокращаться и к месячному возрасту уменьшается в 2 раза. Относительно небольшое по объему мозговое вещество надпочечников, напротив, в первые 6 мес. после рождения увеличивается. У годовалого ребенка каждый из надпочечников весит меньше 1 г. Рост надпочечников продолжается, и у взрослых людей их общая масса достигает 8 г. Примерно к 3-летнему возрасту пучковая и клубочковая зоны полностью дифференцируются, однако сетчатая зона остается недостаточно дифференцированной вплоть до периода полового созревания.

Рост надпочечников на ранних стадиях внутриутробного развития не зависит от АКТГ, но со средины гестационного периода и до его окончания рост и созревание этих желез находятся под контролем АКТГ. Стадия внутриутробного развития, на которой формируется обратная связь между кортизолом и АКТГ, установлена недостаточно точно, но, как свидетельствует клинический опыт, уже в 1 триместре беременности система обратной связи между надпочечниками и гипофизом у плода функционирует нормально. К дополнительным факторам, принимающим участие в регуляции роста надпочечников и стероидогенеза у плода, относятся ХГ и ряд пептидных ростовых факторов, с вырабатываемых плацентой и плодом.
Для развития надпочечников необходимы два фактора транскрипции: SF-1 и DAX-1. Первый участвует также в регуляции транскрипции ряда генов, кодирующих ферменты стероидогенеза. Ген SF-1 расположен на хромосоме 9q33, него разрушение приводит к агенезии надпочечников и гонад, отсутствию гонадотропных клеток гипофиза и недоразвитию вентромедиального гипоталамуса. Мутации гена DAX1, расположенного на хромосоме Хр21, сопровождаются врожденной гипоплазией надпочечников и вторичным гипогонадизмом. DAX-1 также играет важную роль в регуляции стероидогенеза.
Биосинтез надпочечниковых стероидов
Исходным субстратом для биосинтеза всех стероидов служит холестерин. Хотя клетки коры надпочечников способны синтезировать холестерин из ацетата, 80% этого предшественника поставляют в надпочечники липопротеиды плазмы. Расположенные на клеточной поверхности специфические рецепторы ЛПНП связывают эти частицы, затем происходит их интернализация вместе с рецепторами (опосредованный рецепторами эндоцитоз). Холестерин в виде эфиров запасается в особых пузырьках. Под действием специфической гидролазы образуется свободный холестерин, который и используется для синтеза стероидных гормонов.
Скорость-лимитирующим этапом стероидогенеза является перенос холестерина через наружную и внутреннюю мембраны митохондрий. В этом переносе участвует ряд белков, в том числе периферический бензодиазепиновый рецептор и белок острой регуляции стероидогенеза (StAR). Последний обладает очень коротким периодом полувыведения, и тропные факторы (например, АКТГ) быстро индуцируют его синтез. Именно StAR определяет быстрые (от минут до часов) изменения в скорости биосинтеза стероидных гормонов.
На внутренней мембране митохондрий от холестерина отщепляется боковая цепь и образуется прегненолон. Эта реакция катализируется 20,22-десмолазой (P450scc, CYP11А1). Подобно другим ферментам цитохрома Р450, этот фермент представляет собой мембранный гемопротеин с молекулярной массой около 50 кДа. На него поступают электроны от НАДФ-зависимой митохондриальной системы транспорта электронов, состоящей из двух дополнительных белков — адренодоксин-редуктазы (флавопротеин) и адренодоксина (небольшой белок, содержащий негемовое железо). Ферменты Р450 используют электроны и 0-, для гидроксилирования субстрата и образования Н20. В процессе отщепления боковой цепи холестерина последовательно происходят три окислительные реакции. Образующийся в результате прегненолон диффундирует из митохондрий и поступает в эндоплазматический ретикулум. Последующие реакции в разных зонах коркового вещества надпочечников различны.
-
Клубочковая зона.
В эндоплазматическом ретикулуме клеток этой зоны прегненолон превращается в прогестерон под действием 3β-гидроксистероиддегидрогеназы, НАД+-зависимого фермента типа дегидрогеназы короткой цепи. Под действием 21-гидроксилазы стероидов, другого фермента цитохрома Р450, прогестерон превращается в 11-Дезоксикортикостерон. Как и все другие ферменты Р450 эндоплазматического ретикулума, Зβ-гидроксистероиддегидрогеназа использует систему транспорта электронов лишь с одним дополнительным белком — цитохром Р450-редуктазой.
Дезоксикортикостерон затем вновь проникает в митохондрии, где под действием альдостерон-синтазы (еще одного фермента Р450, структурно сходного с 20,22-десмолазой) превращается в альдостерон. Альдостеронсинтаза также осуществляет три последовательные окислительные реакции: 11 p-гидроксилирование, 18-гидроксилирование и дальнейшее окисление С18-метильной группы в альдегид.
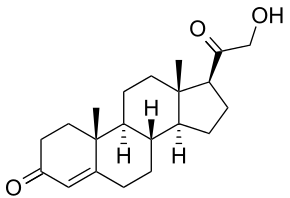
-
Пучковая зона.
В эндоплазматическом ретикулуме клеток пучковой зоны прегненолон и прогестерон под действием 17а-гидроксилазы превращаются соответственно в 17-гидроксипрегненолон и 17-гидроксипрогестерон. В клубочковой зоне 17а-гидроксилаза не экспрессируется, поэтому 17-гидроксилированные стероиды в этой зоне не образуются. В пучковой же зоне 17-гидроксипрегненолон превращается в 17-гидроксипрогестерон и 11-дезоксикортизол под действием тех же ферментов (Зβ-гидроксистероиддегидрогеназы и 21-гидроксилазы), которые функционируют в клубочковой зоне. Поэтому наследственные дефекты этих ферментов сказываются на синтезе и альдостерона, и кортизола. Наконец, 11-дезоксикортизол вновь проникает в митохондрии, где под действием 1ip-гидроксилазы превращается в кортизол. 1 ip-гидроксилаза сходна с альдостеронсинтазой, но обладает меньшей 18-гидроксилазной активностью и полностью лишена 18-оксидазной активности. Поэтому в нормальных условиях альдостерон в пучковой зоне не образуется.
-
Сетчатая зона.
В этой зоне и, до некоторой степени, в пучковой 17-гидроксилаза обладает дополнительной способностью расщеплять связь С17- С20, что превращает 17-гидроксипрегненолон в ДЭА. Под действием Зβ-гидроксистероиддегидрогеназы ДЭА превращается в андростендион, а последний в других тканях — в тестостерон и эстрогены.
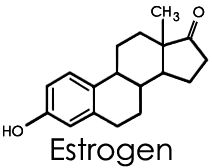
-
Фетоплацентарный комплекс.
В надпочечниках плода синтез стероидов протекает иначе, чем после рождения. Главным источником холестерина для синтеза стероидов в надпочечниках плода служит образующийся в печени холестерин ЛПНП. Активность Зβ-гидроксистероиддегидрогеназы в надпочечниках плода крайне низка, а сульфокиназы стероидов — высока. Поэтому главными стероидными продуктами надпочечников плода являются ДЭА и ДЭА-сульфат. В печени ДЭА-сульфат подвергается 16а-гидроксилированию, превращаясь в 16а-гидроксисульфаз ДЭА. В плаценте, обладающей высокой стероидсульфатазной активностью, из ДЭА и его сульфата образуется эстрон и эстрадиол, а из 16а-гидроксисоединения — эстриол. Для синтеза эстрона и эстрадиола плацента в равной степени использует предшественники из крови матери и плода, предшественники же эстриола поступают почти исключительно от плода. Надпочечники плода вырабатывают не только субстраты для синтеза эстрогенов в плаценте, но и значительные количества кортизола, который под действием 1 ip-гидроксистероиддегидрогеназы превращается в кортизон. К концу внутриутробного периода концентрация кортизола в крови плода возрастает за счет усиления его секреции и меньшего превращения в кортизон. В середине беременности надпочечники плода вырабатывают мало альдостерона, но ближе к рождению его секреция увеличивается.








- Аллергия
- Ангиология
- Болезни глаз
- Венерология
- Гастроэнтерология
- Гинекология
- Дерматология
- Здоровое питание
- Инфекционные болезни
- Кардиология
- Косметология
- Лекарства
- Лекарственные растения
- ЛОР-заболевания
- Мужское здоровье
- Неврология
- Неотложная помощь
- Новости
- Онкология
- Ортопедия
- Паразитология
- Педиатрия
- Пульмонология
- Расшифровка анализов
- Симптомы
- Системные заболевания
- Стоматология
- Травматология
- Урология
- Хирургия
- Эндокринология
- Нужно знать
- Еда
- Профессиональные заболевания
Комментарии